光合作用作为地球上生物利用太阳能的重要反应,一直是科学研究关注的重点,是植物抗逆性研究、作物高产研究的热点。光合作用根据其反应阶段可以分为基于光能吸收传递转化的光反应和基于CO2同化等酶促过程的暗反应。光反应作为植物利用太阳能的原初反应,光能的吸收传递和转化主要发生在植物叶片或者藻类的类囊体膜上,由光系统II(PSII)、细胞色素b6f(Cytb6f)、光系统I(PSI)、ATP合酶(ATP Synthase)共同完成,光反应会产生NADPH和ATP两种用于暗反应的还原剂(同化力),并释放氧气。目前最常用的测量光合作用光反应的手段是叶绿素荧光仪法。近几年随着冷冻电镜技术的发展,有关类囊体膜蛋白超级复合物结构解析的报道越来越多。在国内,中国科学院植物研究所、中国科学院生物物理研究所、清华大学的科研人员在该领域成果卓著,已经成功解析了菠菜PSII–LHCII、豌豆PSI-LHCI、玉米PSI-LHCI&LHCII、水稻PSII-PBCP、红藻PSI-LHCR、绿藻PSII–LHCII、硅藻PSII-LHCII的晶体结构。在国际上,知名的光合作用蛋白复合物结构研究机构有:日本冈山大学理学院生物科学系,嗜热蓝藻OEC和PSII复合体结构的解析均有该单位的科学家完成;德国马克斯·普朗生物理研究所结构生物学系,来自该单位的科学家在2018-2019年成功解析了蓝藻PSI-Ferredoxin和菠菜ATP Synthase的结构;英国谢菲尔德大学的Matthew P. Johnson研究团队则一致专注于Cytb6f的结构,先后在2014年和2019年报道了菠菜Cytb6f结构的研究结果。生理活性是光合作用的外在表现,膜蛋白结构则是生理活性的内在基础。结构的解析可以帮助我们认识光合作用的工作原理,从而更好地利用光合作用解决人类面临的粮食、能源、环境等问题。下面我们按照光合作用光能利用的顺序简单分析一下光合膜蛋白复合体的结构解析带给我们的新启示。

光捕获是光合作用初始的重要过程。在绿色植物中,太阳能收集是由叶绿体类囊体膜中的一系列捕光复合物(色素蛋白)来实现。LHC II是叶绿体中最丰富的整合膜蛋白,以三聚体的形式存在,并与类囊体叶绿素分子的一半结合。每个LHC II单体均包含约232个氨基酸残基的多肽,13-15个Chla和Chlb分子,3-4个类胡萝卜素和一个紧密结合的磷脂。除了光收集功能外,LHC II在强光条件下产生的过量激发能的无辐射消散中也发挥作用。当光强度趋于饱和时,通过激发这种光保护机制,LHC II可以弱化过剩光的破坏效应。此外,LHC II还参与调节光系统II和I的激发能量分布。

2004年3月18日,Nature以封面彩图的形式发表来自中国科学院生物物理所常文瑞课题组题为“Crystal structure of spinach major light harvesting complex at2.72 A ˚ resolution”的研究论文。该研究首次解析了菠菜光合作用系统Ⅱ的主要捕光复合物(LHC-Ⅱ)的晶体结构。结果发现大型R32晶胞的一个不对称晶胞包含十个LHC-II单体。每个单体中的14个叶绿素(Chl)可以明确区分为八个Chla和六个Chlb分子。每个叶绿素的跃迁偶极矩的方向分配已经实现。所有Chlb都位于相邻单体之间的界面周围,并且与Chla一起是高效采光的基础。每个单体有四个类胡萝卜素结合位点。单体-单体界面处的叶黄素循环类胡萝卜素可能参与了多余能量的非辐射消散,这是植物中发展起来的一种光保护策略。

LHC-II三聚体和单体中的色素布局,立体视图显示了LHC-II 三聚体中的色素排列模式。单体标记为I-III。为清楚起见,省略了叶绿素植物链和脂质。绿色,叶绿素a;蓝色,叶绿素b;黄色,叶黄素;橙色,新叶黄素;品红,叶黄素循环类胡萝卜素。b,c,分别为基质侧和腔侧观察的单体中的色素图像。颜色名称与a, d, e相同,分别在基质侧和腔侧的LHC-II三聚体内叶绿素的排列。叶绿素由三个原子表示:中心镁原子和两个氮原子。两个氮原子之间的连线决定了量子跃迁偶极矩的方向。绿色,CHLA氮;蓝色,CHLB氮;灰色,镁;紫色和蓝色椭圆形,近似单体面积。连接两个叶绿素的暗线附近的洋红色数字表示它们之间的中心到中心的距离。

基于结构的非光化学猝灭(NPQ)模型。从基质侧沿二十面体C2轴的顶视图。 DGDG显示为黄色透明的球棒模型。叶绿素和叶黄素循环类胡萝卜素的代表如前图所示。黑色箭头表示从一个三聚体到相邻三聚体的激发能转移途径,橙色箭头显示从叶绿素Qy到叶黄素循环类胡萝卜素S1的可能转移途径。红色星号表示推定的淬灭位点。为了清楚起见,一个三聚体中的字符为黑色,而另一个三聚体中的字符为灰色。
PSII的捕光天线是一个复杂的复合物,LHC II因其在叶绿体中的丰度很高而被称为主要的捕光复合物,位于PSII的最外围。除此之外还有其他三个单体捕光复合物,分别称为CP29,CP26和CP24,位于LHCII和核心复合物之间。这些是次要的捕光复合物,与LHCII相比,它们叶绿素含量相对较低。它们除了在收集太阳能并将其转移到反应中心方面发挥作用外,这些内周天线复合物还可以在高光照条件下耗散多余的激发能。这种机制通常称为非光化学猝灭(NPQ)。

2011年2月6日,Nature Structural & Molecular Biology杂志在线发表了中国科学院生物物理研究所常文瑞院士课题组题为“Structural insights into energy regulation of light-harvesting complex CP29 from spinach”研究论文。该研究解析了菠菜次要捕光复合物CP29的2.8分辨率晶体结构,并在晶体结构的基础上深入分析讨论了CP29的捕光、能量传递和光保护等功能。这是国际上首个高等植物次要捕光复合物的晶体结构。
研究发现每个CP29单体包含13个叶绿素和3个类胡萝卜素分子,这与主要的捕光复合物LHC II有很大不同。将这13个叶绿素结合位点分配为8个叶绿素a,4个叶绿素b和1个被叶绿素a和b占据的假定混合位点。基于X射线结构分析,在CP29中构建了一个集成的色素网络。确定为两个特殊的色素分子簇,分别是a615-a611-a612-Lut和Vio(Zea)-a603-a609,它们可能充当潜在的能量猝灭中心以及能量转移途径的出入口。
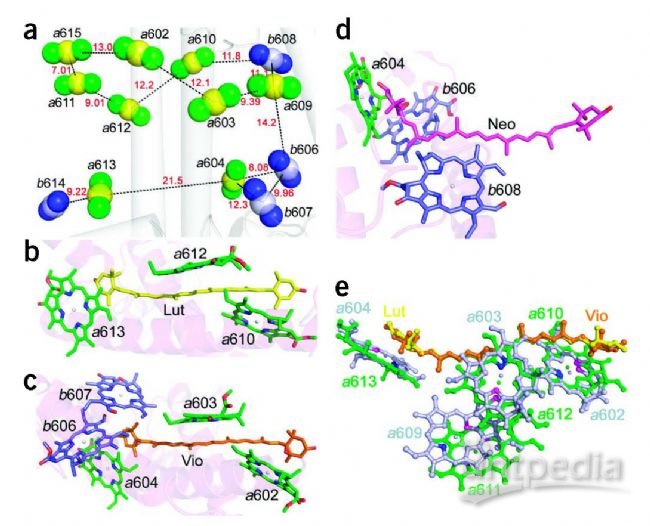
CP29中的色素排列。(a)沿膜平面视图的Chls分布。Chls由三个原子表示:中心镁原子和两个氮原子(NB,ND)。黄色:叶绿素a镁;白色,叶绿素b镁;绿色,叶绿素a氮;蓝色,叶绿素 b氮。每层中相邻的Chls,两层中最接近的Chls和腔层中两个簇(Chls a604-b606-b607和Chls a613-b614)中最接近的Chls用黑色虚线连接。它们的中心距离(Å)用红色数字标记。Chls分子中心的坐标是通过四个吡咯氮原子的坐标算数平均来计算的。(b–d)Lut,Vio,Neo及其附近的Chls。(e)Lut和Vio的叠加显示出与相邻的Chls类似的布局。绿色,在Lut附近灰色,在Vio附近。每个Chla中的氮原子NB和ND分别以蓝色和品红色表示。

强相互作用的色素团a615–a611–a612–Lut。(a,b)a611–a615的Chl对。氢键用暗虚线相连。Lys199与G3P之间的离子键用蓝色虚线(a)表示。Chls a611和a615的Qy跃迁偶极矩分别用绿色和青色箭头标记(b)。(c)色素簇a615–a611–a612–Lut的立体视图。连接两种色素的黑色虚线附近的数字表示色素之间的中心距离。
从LHC
II三聚体和CP29单体结构的解析我们可以得到一些启示,CP29和LHCII的结构差异,特别是它们的色素组成和空间排列,可能是它们在光合捕光和调控过程中功能不同的原因。在CP29结构中新发现的Chl
a615与Chls
a611-a612和Lut一起,组成了一个强相互作用的色素团簇,可能是能量猝灭中心的潜在位点。在过剩光照条件下,类囊体腔中由低pH(酸化)引发的叶黄素循环(紫黄质通过花药黄质脱环氧化成玉米黄质(Zea))被认为是NPQ中的一个重要途径。在体内CP29中观察到Vio向Zea的转化效率相对较低,但在强光条件下和长时间暴露在光下时,Vio向Zea脱环氧化的程度增加。此外,低pH值下酸性残基质子化引起的CP29构象变化对NPQ也有重要意义。
PSII的外围天线是一个高度复杂的系统,具有很大的复杂性和多功能性。三聚体LHCII与单体CP29、CP26和CP24共同承担着收集太阳能和光保护的重要功能。他们既相互合作,又有独立的角色。这反映在它们在整体结构上有着明显的相似性,却在色素组成和排列上有着实质性的差异。CP29的三维结构为我们进一步了解PSII的小型外围天线提供了机会。我们相信,当CP26和CP24的三维结构可用时,特别是当获得超复杂体的整体结构(包括所有这些外围天线)时,我们将更全面地了解PSII外围天线的功能机制。

除了NPQ型非光化学淬灭之外,还存在一种qE型的非光化学淬灭,光系统II蛋白PsbS在qE型非光化学猝灭中起重要作用,可保护植物免受过量光照条件下的光损害。qE是由低pH值激活PsbS引发的。但是PsbS作用的机制一直不清楚。直到2015年8月10日,Nature Structural &Molecular Biology在线发表了中国科学院生物物理研究所常文瑞院士/李梅研究组题为“Crystal structures of the PsbS protein essential for photoprotection in plants”的研究论文。论文报告了菠菜中低pH下游离形式的PsbS的晶体结构,以及与qE抑制剂N,N'-二环己基碳二亚胺(DCCD)形成复合物,揭示了PsbS采用独特的折叠模式,这不同于其他的捕光复合物超家族,它是一种非经典的色素结合蛋白。结构和生化证据表明,活性和非活性PsbS在类囊体膜中均形成同型二聚体,DCCD的结合破坏了活性PsbS二聚体的腔内分子间氢键。在qE期间通过低pH值激活PsbS可能涉及与PsbS二聚体的腔内分子间相互作用改变相关的构象变化相关。本研究成果从结构和生化分析为PsbS的活化和抑制机制提供了分子基础。然而,激活的PsbS如何启动qE尚待阐明。
已有的研究表明,在强光条件下,最初与PSII核心相关的PsbS向LHCs28迁移。如果激活的二聚体PsbS蛋白被插入到触角之间,而没有内部色素分子来传递激发能量,它可能会作为衰减器来显著减缓LHCs之间的能量流动。此外,激活的PsbS二聚体可能直接与相邻的LHCs相互作用,促进其构象变化,并在LHCs内部或接触界面诱导淬灭。之前有研究人员成功的用含有PsbS、LHCII和玉米黄质的蛋白质脂质体系统体外重组淬灭过程支持了PsbS和LHC在qE期间的直接相互作用。PsbS也有可能仅仅作为pH传感器或催化剂间接参与qE。比本研究稍早的研究表明,qE的形成与类囊体膜中PSII-LHCII超复合物的结构重组有关,并表明PsbS控制着这种重组,这可能是由低pH诱导的PsbS二聚体构象变化引起的。这里提出的PsbS结构为进一步研究PsbS在qE中的作用机理奠定了基础。

在光合作用过程中,植物光系统II核心复合体接收来自外围捕光复合体II(LHCII)的激发能。最初的光物理和光化学过程都是由光系统II(PSII)完成。PSII是一种嵌入类囊体膜内的超分子复合物。它包含许多蛋白质亚基和各种辅助因子,包括叶绿素、类胡萝卜素、一个Mn4CaO5簇、一个血红素、质体醌和脂质体。PSII的一个特征功能特性是它通过Mn4CaO5团簇催化的光诱导水氧化反应从水分子中提取电子的能力。为了收集光子能量并驱动光化学反应,植物PSII含有一系列的外围捕光复合物(主要的捕光复合物是PSII(LHCII),次要的捕光复合物叶绿素结合蛋白(CP29、CP26和CP24))。这些天线复合物围绕PSII的核心复合物,吸收光能并将其传输到反应中心,以诱导称为P680的特殊叶绿素对中的电荷分离。然而,由于对PSII-LHCII超复合物高分辨率的结构缺乏认识,植物PSII的外周天线与核心复合物之间激发能量传递的精确途径仍不清楚。此外,植物PSII中三个重要的外源亚基(PsbO、PsbP和PsbQ)的功能和相互作用尚不清楚。为此中科院生物物理所柳振峰团队通过单颗粒冷冻电镜技术,在3.2 Å分辨率下解析了1.1兆道尔顿的菠菜光系统II–LHCII超级复合物的结构。首次揭示了LHCII、CP29以及CP26向核心天线复合物CP43或CP47传递能量的途径,对于进一步在分子水平理解PSII-LHCII超级复合物中的能量传递时间动力学和光保护机理具有重要意义。相关结果于2016年5月18日发表在Nature杂志上,题目为 “Structure of spinach photosystem II–LHCII supercomplex at 3.2Å resolution”,电镜结果显示PSII-LHCII是一个同源二聚体的超分子体系,其中每个单体含有25个蛋白亚基,105个叶绿素分子,28个类胡萝卜素分子和其他辅因子。三个外部亚基(PsbO, PsbP 和 PsbQ)对于光系统II的最佳放氧活性至关重要,它们形成了一个三角形冠状结构,环绕着CP43和D1蛋白的Mn4CaO5结合结构域(光系统II中的1个Ca原子与4个 Mn原子由5个O原子作为桥连成Mn4CaO5簇,是光催化水分子断裂生成氧气的放氧复合体的核心)。一个主要的三聚体LHCII和两个次要的单体LHCII分别与核心复合物的每一个单体结合,且三个小的内部亚基(PsbW, PsbH 和 PsbZ)加强了天线与核心复合物间的相互作用。通过分析紧密连接的界面间叶绿素分子,我们对天线与核心复合物间的能量传递通路有了更深入的了解。

菠菜PSII-LHCII超复合物的总体结构。a,b,菠菜C2S2型PSII-LHCII超复合物的结构。a,沿膜从基质侧水平角度观察。b,沿膜垂直角度的侧视图。虚线表示两种单体之间的估计界面区域。主要组件显示为不同颜色的卡通和棒状模型,而12个小的内在亚基显示为黄色球形模型。c,卡通图,显示了超复合物的膜嵌入亚基的分配。仅显示一种单体,并且颜色代码与a中一致。d,超复合体的腔侧暴露区域。该视图是沿腔从腔侧面观察的。

PSII-LHCII超复合物中的天线-核心和天线-天线相互作用。a,LHCII和CP43 / D1之间的接口。它们的相互作用由PsbW和PsbI介导。 PsbW(CTR),PsbW亚基的C端区域。b,CP29和CP47 / D1之间的相互作用。 M4–5环,D1的第四个和第五个跨膜螺旋之间的环区域;M3和M4,分别是CP47的第三和第四跨膜螺旋;M2–3环,CP47的第二个和第三个跨膜螺旋之间的环;PsbH(NTR),PsbH亚基的N端区域。c,CP26和CP43 / PsbZ之间的相互作用。d,LHCII和CP29'之间的接触。e,CP26–LHCII相互作用。

从天线复合体到反应中心的能量转移途径。a,PSII-LHCII超复合物中叶绿素的分布模式。Chla和Chl b分别用绿色和蓝色区分。箭头指示LHCII和CP29'/ CP26(红色),LHCII / CP29'/ CP26和核心天线(CP43 / CP47')(洋红色),CP43 / CP47'和D1 / D2'反应中心之间的潜在能量转移路径(青色)。五个红色椭圆形表示在CP29'中Chla611-Chla612对周围的潜在能量猝灭位点,黑色椭圆形表示在CP29'中Chla603-Chla609对周围的另一个潜在淬灭位点。这些淬灭位点可能在强光条件下被激活,以消散有害的过剩能量。虚线表示每个单独复合物的近似边界。 b–d,界面叶绿素支持LHCII和CP29'(b),LHCII和CP26(c)以及LHCII和CP43(d)之间的能量转移。 e,f,CP29和CP47(e)之间以及CP26和CP43(f)之间的界面处的叶绿素。虚线附近的数字表示两个相邻叶绿素之间的Mg至Mg距离(Å)。界面叶绿素以洋红色突出显示。
综上所述,我们可以深入了解PSII中的能量传递途径。通过位于周围和核心天线复合体之间的界面叶绿素对,激发能可以从LHCII,CP26和CP29传递到CP43或CP47。LHCII三聚体在其单体-单体界面处包含三个富含Chb的簇,其中两个簇连接到CP26和CP29'。由于Chlb的能量水平高于Chla,LHCII和CP26之间(或LHCII和CP29'之间)的能量转移可能会从LHCII的富含Chlb的区域流到CP26(或CP29')中与之相邻的富含Chla的区域。对于LHCII和CP29'之间的能量传递,来自腔侧面的Chl b605monomerA/LHCII和Chl a604 / b606CP29'形成最接近的内复合物对,Mg-Mg距离(Dcentre)为17.7 / 18.6Å,而在基质层Chlb608monomerA/LHCII和Chlb608CP29'连接的Dcentre为23.0Å。因此,CP29'的富含Chlb的区域与LHCII的区域相连,促进了二聚体超复合物的两个相邻单体之间的能量转移,大概是通过Chlb605monomerA/LHCII 到 Chla604 / b606CP29'和b608monomerA / LHCII到b608CP29'的路径。对于LHCII和CP26之间的能量传递,来自LHCII三聚体的单体B的Chlb608连接到CP26的Chla612和Chla610,连接Dcentre分别为21.8和21.9Å,而来自腔侧面的LHCII的Chlb605可能转移其激发能量到CP26的Chla604中,因为它们的连接Dcentre为19.3Å。
LHCII中最低能态的叶绿素为Chl
a610,a612和a611簇,被称为末端发射域。LHCII三聚体中平衡的激发能将集中在该簇上。在超复合物中,来自LHCII三聚体的单体A的末端发射体可能会将其能量传输到Chla506CP43,该基团相对于Chla611monomerA/LHCII位于有利的方向(几乎平行)和距离(17.1Å
Dcentre)。在Chla611下方,来自腔侧面的LHCII的Chla614以25.1Å的Dcentre连接到Chla501CP43。这两个途径构成了LHCII和核心复合物之间能量转移的基础。在没有次要天线复合体的情况下,LHCII可以将能量直接转移至核心复合体,但是在次要天线敲除的突变体植物中,LHCII和PSII核心之间的功能连接受到严重损害。
为了在CP29和CP47之间进行能量转移,将Ch1a616CP29夹在Ch1a609CP29和Ch1a616CP47之间,Dcentre分别为9.3 Å和14.4 Å。Ch1a616CP29和Ch1a609CP29 / a616CP47之间最接近的边对边距离(Dedge)为3.4 / 4.2Å,表明这些叶绿素形成了强耦合对。Ch1a616CP29在间隙中的位置使其成为关键的连接子,将激发能从CP29传递到CP47。此外,CP29的Chla603是以18.6Å 的Dcentre直接连接到Chla610CP47的。在腔侧面,能量可能会以19.1 Å 的Dcentre从Ch1b607CP29转移到Ch1a607CP47。或者Ch1b607CP29将它的能量传递给Ch1a603-a609CP29,然后能量进一步由Chl a616CP29 释放给 Chl a616CP47。在这些潜在途径中,Ch1a616CP29至Ch1a616CP47可能是CP29和CP47之间最有效的能量转移途径,因为这两个叶绿素在界面上配对最紧密。
CP26与CP43相互作用密切,它们之间的能量转移可能通过多种潜在途径发生。Ch1a611CP26与最红的Ch1a612形成一个强耦合对,该耦合对可能是CP26的终端发射域。Chla611以18.2Å 的Dcentre连接到Ch1a512CP43,而以17.2Å 的Dcentre连接到Chla513CP43。同时,Chlb601CP26以4.8 Å 的Dedge(Dcentre为12.6 Å)与Chla513CP43耦合,并且还以19.2 Å 的Dcentre连接到Chla512CP43。在腔侧面,来自Ch1a614CP26的激发能量可能主要被16.0 ÅDcentre 处的Ch1a503CP43吸收。因此,从CP26传输的能量将在基质侧被Chla513–Chla512接收,或者被CP43腔侧面的Chla503接收。
当外围天线复合体的激发能被核心天线复合体收集,随后的能量通过位于CP43,CP47,D1和D2内的Chla网络从CP47或CP43传递到P680特殊叶绿素分子对。在强光条件下,主要和次要LHCII中的色素分子簇可充当非光化学猝灭位点,将有害的多余激发能耗散为热量。超复合体内的潜在猝灭位点主要位于相邻天线复合体之间的界面处或附近。这些位置非常适合它们在到达反应中心之前拦截和消耗多余的能量。比该研究稍早一些时候的报道显示,生物物理模型研究已经获得了有关PSII-LHCII超复合物中光收集动力学的初步信息。现在,菠菜PSII-LHCII超复合物的冷冻电镜结构为其高度复杂的色素网络提供了详细的框架,并使人们能够更深入地了解超复合物中光捕获过程的动力学和调控。
由于相关研究内容非常专业,难免有些理解不准确或者编辑整理的疏漏,请以英文原文为准。
参考文献
1. Liu Z, Yan H, Wang K, et al. Crystal structureof spinach major light-harvesting complex at 2.72 Å resolution. Nature,2004, 428(6980): 287.
2. Pan X, Li M, Wan T, et al. Structural insights into energy regulation of light-harvesting complex CP29 from spinach. Nature structural & molecular biology, 2011, 18(3): 309.
3. Fan M, Li M, Liu Z, et al. Crystal structures of the PsbS protein essential for photoprotection in plants. Nature structural & molecular biology, 2015, 22(9): 729.
4. Wei X, Su X, Cao P, et al. Structure of spinach photosystem II–LHCII supercomplex at 3.2 Å resolution. Nature, 2016, 534(7605): 69.




 首页
首页


