DGE分析是使用RNA-seq来检测稳态下的mRNA表达水平,这一表达水平是通过mRNA的转录,加工和降解速度来决定的。但是,RNA-seq也可以用于研究涉及转录,翻译所涉及的过程与动力学特征,这些研究为基因表达提供了新的思路。
基因表达是一个内在的动态过程,但是在检测复杂转录应答的细微以及快速变化或确定不稳定的非编码RNAs,例如增强子RNAs方面,常规的DGE分析方法就比较受限。RNA-seq可以用于绘制TSSs以及定量新合成的新生RNA,这就可以用来研究RNA动力学。但是,与DGE分析相比,nascent RNA的分析则比较难,因为它们半衰期短,丰度低。因此,为了研究这些动态的重要性,研究者们就开发了多种方法来分析nascent RNA;这些方法揭示了在启动子处的差异转录程度,表明RNA聚合酶II(Pol II)在启动子附近的暂停是基因表达的关键调节步骤,证明了nascent RNA有直接调节转录的作用,并表明其序列和结构影响转录的延伸,暂停和停顿,以及发挥染色体修饰结合和增强了子的作用。nascent RNA- seq方法旨在区分新近转录的RNA和其它RNAs,这些方法可以分为3类:run-on方法,Pol II免疫沉淀法,代谢标记法(FIG. 4)。
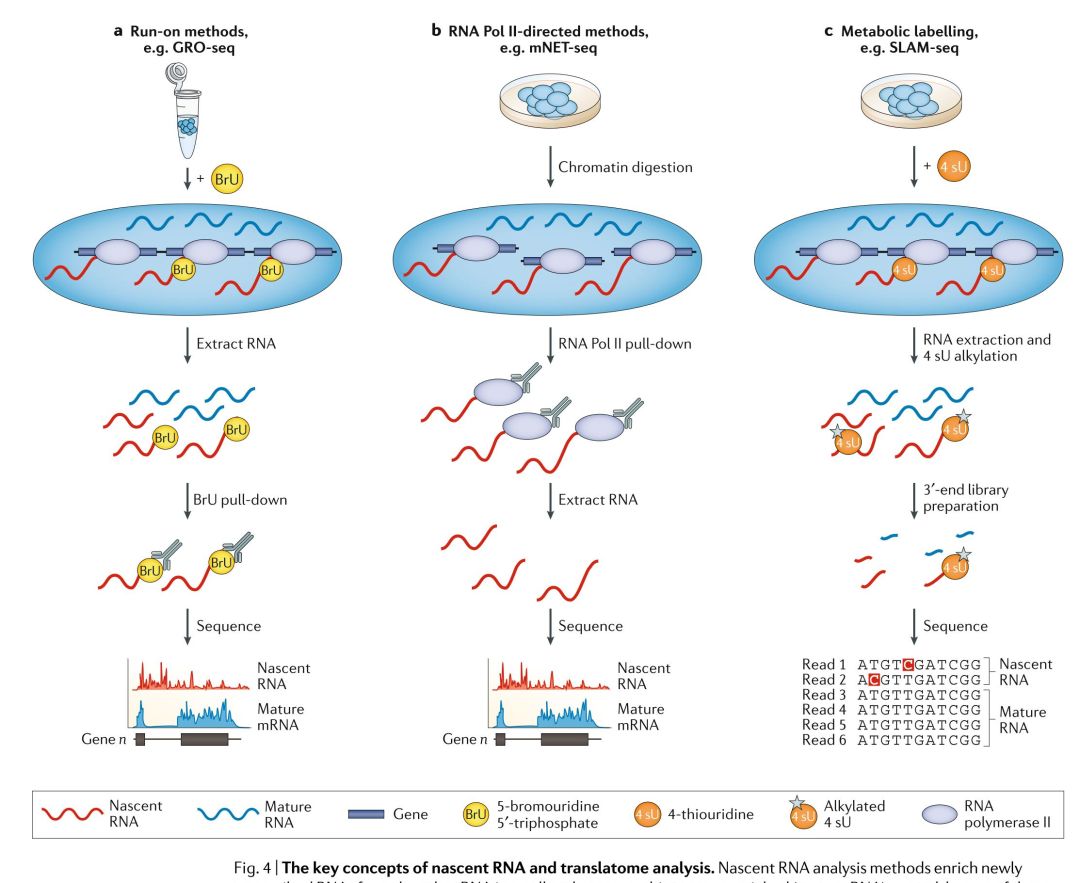
Run-on法是将核酸类似物添加到样品中,从而使nascent RNA能够从总的RNA混合物中进行富集,并能够检测瞬时RNA的转录(FIG. 4a)。全局run-on测序(Global run-on sequencing, GRO-seq)与精确核酸run-on测序(Precision nuclear run-on sequencing, PRO-seq)是分别将Bru或生物素修饰的核酸在RNA的转录期整合到nascent RNA中来实现的。其过程大致为,分离细胞核,并通过洗涤除去内源性核苷酸,再添加外源生物素标记的核苷酸,随后恢复转录。通过免疫沉淀或亲和纯化的方法,对富集的新转录RNA进行测序,从而检测参与转录的RNA聚合酶的位置和活性。由于run-on过程中标记的核苷酸的数据,GRO-seq只能测到10-50bp的长度,这就降低的TSS检测的精度。PRO-seq能够实现单个碱基级的分辨率,因为生物素标记的核苷酸掺入后转录就停止,可以识别出转录位点。Run-on方法理解起来很简单,就是RNA分子整合了修饰的核苷酸,并对其进行富集,用于测序,但是在实践中,背景中存在有non-nascent RNA,这就需要增加读长深度。利用这些方法,提示了启动子处,启动子处差异或双向转录本起始的程度,确定了增强子RNA在调节基因表达方面的作用。通过特定富集5’加帽的RNAs,GRO-cap,PRO-cap或small 5’capped RNA测序(small 5ʹ-capped RNA sequencing, START-seq)增加了检测转录起始和捕获RNAs的灵敏度和特异性,这种处理还会降低源于转录后加帽的RNAs的背景信号。
Pol II的免疫共沉淀方法包括,天然延长转录测序(native elongating transcription sequencing, NET-seq)和哺乳动物染色质天然转录测序法(native elongating transcript sequencing for mammalian chromatin, mNET-seq),使用抗FLAG(用FLAG标记的Pol II)抗体进行沉淀的方法,或各种针对Pol II C末端结构域(CTD)的沉淀方法(FIG. 4b)。与这些染色质复合物结合的nascent RNA的RNA-seq方法用于检测TSSs,虽然non-nascent Pol II结合的RNA与背景mRNA会对读长浓度产生负面影响,影响分析。NET-seq缺乏特异性,因为任何与Pol II强烈结合的RNA都会污染nascent RNA的富集效果,例如在NET-seq数据中就存在有tRNA和small nucleolar RNA。在mNRET-seq中使用多个CTD抗体提示了VTD修饰是如何影响转录的,检测到了RNA加工的中间体,并能能够将特定的Pol II nascent RNAs定位于TSSs。然而,这些检测能力是以更复杂的实验,更多的细胞数量和更高的测序成本为代价的。
使用核苷酸类似物硫代吡啶(4-thiouridine, 4 sU)进行代谢脉冲标记(Metabolic pulse- labelling)的方法可以识别nascent RNA(FIG. 4c)。但是,在那些需要长标记时间的方法中,大多数的转录本都会被标记,这就限制了这种方法的灵敏度。通过专门针对RNAs的3’末端(仅最近拉RNA聚合酶的新转录的RNA)的方法,瞬时转录组测序(transient transcriptome sequence, TT-seq)与硫醇(SH, thiol)连接的烷基化RNA代谢测序(thiol(SH)-linked alkylation for metabolic sequencing of RNA, SLAM-seq)能够降低来源于5’RNA的信号。TT-seq将标记时间限制在5分钟,因此只标记新转录本的3’末端,它在进行生物素亲和纯化前,有一个RNA片段化操作,用于富集标记的RNA。SLAM-seq整合了3’mRNA-seq文库制备方法(虽然它也用于其它的文库制备,例如miRNA), 它仅针对标记的新转录的RNA进行测序,而非整个转录本进行测序。此外,在SLAM-seq中,提取RNA后,还要加入碘乙酰胺(iodoacetamide),用于烷基化已经插入到新生成的nascent RNA链中的4 sU残基。这种修饰会诱导反转录式依赖的胞腺嘧啶到胞嘧啶的转换(T > C),这在测序分析中会被检测为“突变”,从而直接识别为4 su整合位点。然而,低掺入率意味着只有少量的4 sU位点可以被转换为胞嘧啶,这就限制了灵敏性。有两种方法,即TUC-seq与TimeLapse-seq也使用T>C这种突变分析方法,但是它们并不富集3’末端。这两种方法用于研究细胞干扰后的转录应答和RNA的半衰期。
Nascent RNA分析方法还未进行过直接比较。Nascent RNA方法都受到非特异性背景和/或降解的RNA的负面影响,这会影响读取深度。通过仅测序3’末端,那么non-nascent RNA的效应就会在PRO-seq,TT-seq和SLAM-seq中降低,但是几乎没有证据表明是否有其他方法更优。亲和纯化方法费时费力,与代谢标记法相比,前者需要更多的起始材料,但是,确定脉冲标记的时间比较复杂,并且短脉冲产生用于分析的RNA很少,这限制了灵敏度。最近开发的,组织特异性RNA标记方法以及亲折突变分析计算方法或许能够促进研究者转向使用生化(基于生物素)富集的手段来研究富含生物学意义的nascent RNA和其它RNA。Nascent RNA方法以及它们与其它方法的隧和,例如空间转录组学或RNA-RNA与RNA-蛋白质相互作用的方法,将会提高我们对转录过程的理解。
 首页
首页






