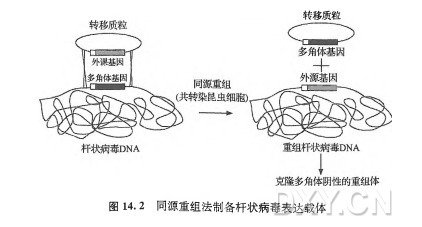
| 实验步骤 | 一、杆状病毒表达载体最简单的经典杆状病毒表达载体是一个重组的杆状病毒,其基因组含有一段外源核酸序列,通常为编码目标蛋白质的dDNA,在多角体蛋白启动子控制下进行转录。这个嵌合的基因由多角体蛋白启动子和外源蛋白编码序列组成,其位于病毒基因组多角体座位,替代了非必需的野生型多角体基因。在实验室中,重组病毒能够感染昆虫细胞或幼虫(毛虫),这会使外源cDNA在感染的极晚期出现高水平的转录。转录出的mRNA经翻译产生目标蛋白质。多角体蛋白启动子似乎总是可以在转录水平上驱动极高水平的外源基因表达,在很多时候可以获得大量的外源蛋白, 就像最初推测杆状病毒能够制造出大量的多角体蛋白那样。确实是这样,具有高水平表达重组蛋白的潜能是杆状病毒-昆虫细胞系统的一个主要优点。在此, 高水平的含义非常宽泛,是指每升感染杆状病毒的昆虫细胞培养物或4 g昆虫细胞(通常细胞密度为 1X106 个细胞/mL)产出大于或等于100 mg的重组蛋白。此外,在杆状病毒-昆虫细胞系统中高水平表达的蛋白质很少形成包涵体,而包涵体常常在细菌表达系统中形成。任何从事重组蛋白生产的研究人员都清楚,在任何表达系统中蛋白质可以达到的产量和可溶水平很大程度上取决于所研究的蛋白质本身。因此, 从25年的杆状病毒-昆虫细胞表达系统使用经验得出的结论是: 与分泌蛋白相比,该表达系统更易获得细胞核蛋白和细胞质蛋白的高表达。前者的表达水平较后者要低, 通常只有每升几毫克到十几毫克 (Jarvis,1997)。 杆状病毒-昆虫细胞系统的另一个优点是具有真核蛋白的加工能力, 包括对蛋白质进行磷酸化或糖基化等的修饰能力。但是,这种观点有一定的局限,现在已经很清楚地认识到,杆状病毒的鱗翅类昆虫细胞宿主的蛋白质加工与更高等的真核细胞不同。另外的困扰是杆状病毒的感染对宿主蛋白质加工有负面的影响(Azzouzetal.,2000;JarvisandSummers,1989)。显然,对于任何有兴趣生产需要进行真核修饰的重组蛋白的研究者来说,特别是已知这种修饰直接或间接的影响功能,这些都是需要重点考虑的因素。 最后指出的是,杆状病毒-昆虫细胞系统已被证明对多蛋白质亚基复合物(multiproteinsubunitcomplexe) 的生产非常有用[参考Berger等 (2004);Kost等(2005) 的综述]。 该系统能够产生由多组分构成的病毒颗粒,这样的病毒颗粒是极好的候选疫苗,这体现了该系统在此重要应用领域的强大功能。例如,杆状病毒-昆虫细胞系统已经用来生产由脊髓灰质炎病毒(poliovirus)、蓝舌病毒(bluetonguevirus)、腺相关病毒(adeno-associatedvirus)、丙型肝炎病毒 (hepatitisCvirus) 和乳头瘤病毒(papillomavirus)来源的多种蛋白质组成的病毒样颗粒。该病毒样颗粒的产生可以通过构建多个表达单一蛋白质的重组病毒来实现,或者构建一个能够编码多个重组蛋白的单一病毒去感染昆虫细胞。 二、杆状病毒表达载体技术一早期如前所述,利用基本的同源重组原理,构建了第一个重组的杆状病毒载体用于表达由多角体蛋白启动子和外源编码序列组成的嵌合基因。该方法的详细技术可以参考本书的第一版(Bradley,1990), 也可以参考原作者发表的主要论文及其编写的优秀的技术手册(O’reillyetal.,1992;SummersandSmith,1987)。因此,和上面一’样,这部分在此不再赞述。但是,为了对背景有所了解, 对此进行简短地介绍还是很重要的。该常用的方法涉及: ①构建具有嵌合基因的细菌转移质粒,该嵌合基因具有来源于病毒基因组多角体区域的侧翼序列(图I4.1); ②使用转移质粒与从纯化的野生型AcMNPV提取的基因组 DNA 的混合物共转染培养的昆虫细胞(图14.2)。 在共转染的昆虫细胞中,转移质粒与AcMNPV基因组通过同源重组产生重组病毒 DNA 分子,该同源重组发生在转移质粒上的目的嵌合基因侧翼序列与病毒基因组多角体蛋白基因的上游和下游序列之间。为了在敲除(knockout)多角体蛋白基因的同时敲人(knock-in) 编码目标蛋白质的嵌合基因,这必须是一个发生了双交换的重组(doublecrossoverrecombination)。当然,这种重组的发生率相对较低,估计在 0.1% 左右(Smithetal.,1983a)。因此, 必须将少数重组病毒的子代病毒从共转染产生的大量的亲代病毒的子代病毒中分离出来. 通过杆状病毒空斑实验可以很容易地完成分离, 但是研究人员要能够从大量的亲代病毒产生的空斑中辨别出重组病毒形成的空斑。最初,可以通过简单的肉眼筛选来进行辨别, 因为来自亲本的子代病毒形成的为多角体蛋白阳性的空斑, 而重组体子代病毒由于缺少表达多角体蛋白的基因,所以呈现多角体蛋白阴性的空斑。经过专业培训的研究人员能够在解剖显微镜下通过观察实验平板鉴定出多角体蛋白阴性的空斑。因此, 有这种辨别能力是筛选成功的关键。多年来,许多研究人员不具有识别多角体蛋白阴性的杆状病毒空斑的能力,这是一个非常严重的问题,它制约着杆状病毒-昆虫细胞系统作为表达重组蛋白生产工具的应用。 三、杆状病毒表达载体技术—-改进从1卯0年左右开始,研究人员开始克服了杆状病毒载体分离方面的技术局限,并且通过对上述基本方法的大量改动,在别的方面对该系统也进行了改进。这些改进可以分为两类: 一类是转移质粒的改进; 另一类是亲代杆状病毒基因组的改进。下面对它们分别进行介绍。 四、杆状病毒转移质粒的改进对转移质粒进行了改进是出于两个不同的目的。最重要的目的是使通过肉眼筛选鉴定重组杆状病毒空斑变得简单,从上面描述的原因可知这曾经很难。通常的一种此类改进方法是引人在杆状病毒启动子控制下的嵌合标记基因。例如,大肠杆菌的/3-半乳糖苷酶蛋白与多角体阴性空斑表型相比,对它的肉眼识别更加容易(Vialardetal.,1990)。 在转移质粒中引人标记基因是聪明的做法,但也应该注意到该种改进方法存在的潜在问题(O’reillyetal.,1992)。弓丨人的标记基因不仅能够作为所需要的双交换同源重组的信号,也能够作为在转移质粒和病毒 DNA之间产生的单交换同源重组(singlecrossoverrecombination) 的信号。单交换重组的发生概率很高,所产生的重组病毒的基因组包含了细菌复制子在内的转移质粒的全部序列,而且整合位点有一定的随机性。这些重组体在遗传上是不稳定的, 新获得的外源基因可能会在 1~2轮的病毒复制中丢失。尽管有很大的局限性, 能将标记基因引入重组病毒基因组的转移质粒还是被广泛使用, 对于那些清楚知道潜在的单交换重组陷阱的研究人员来说, 它们尤其有用。他们只是使用标记基因进行初筛, 然后通过进一步筛选来确定外源基因在基因组中的整合位点,最后确定是否是双交换重组将目的基因转移到了重组杆状病毒载体。 对转移质粒进行改进的第二个目的是方便重组蛋白在杆状病毒-昆虫细胞系统的表达与纯化。这种改进的数量太多因而不能在此逐一讨论, 有3 种通常的改进属于此类,它们包括引人信号肽的编码序列; 在蛋白质的N端或C端引入纯化标签的序列(如His6); 将多角体蛋白启动子替换为别的杆状病毒启动子, 或者使用多个启动子元件替换多角体蛋白启动子,这样就可以在杆状病毒感染时同时表达多个重组体蛋白。 最近的一个出版物列出了几乎所有的经过上述两类改进的杆状病毒转移质粒,并且简短描述了它们功能上的特色,它是该领域一个很好的信息来源(PosseeandKing,2007)。但是,有一类改进并未列出但值得进一步的关注,它们是即刻早期(immediate^early)转移质粒,在这些质粒中,多角体启动子被替换为杆状病毒即刻早期基因的启动子,如AcMNPV的iel基因 (Jarvisetal.,1996)。使用来自杆状病毒早期基因的启动子 (如iel基因)似乎违反之前提及的原则,因为这些启动子比多角体蛋白启动子弱。但是,有一些证据可以表明,在早期启动子控制下重组杆状病毒表达外源基因可以获得更高质量的蛋白质产物,尽管这些早期启动子不能启动同样高水平的转录(ChazenbalkandRapoport,1995;Hill-PerkinsandPossee,1990;Jarvisetal.,1996;Murphyetal.,1990;Rankletal.,1994;SridharetaUW93)。事实上,这个方法尤其适用于分泌蛋白。如上所述,这些蛋白质在多角体蛋白启动子控制下通常是低水平的表达。在这种情况下, 研究人员或许能够利用这种在感染较早期进行外源基因表达的好处, 从而避免杆状病毒感染对宿主蛋白加工途径带来的负面作用,同时也能够获得高水平表达的产物。 |
|---|








 首页
首页