抗体,一方面是免疫诊断试剂研发的原料,另一方面是免疫诊断试剂检测的对象。曾经以为从学生时代开始就接触抗体,应该对抗体了解的十分透彻,但最近在工作中发现,其实抗体并不是那么简单。所以,翻出免疫学教材,对抗体做一番回顾和学习。
抗体分为分泌性免疫球蛋白(血清、体液及分泌液)和膜免疫球蛋白(mIg,BCR),IVD行业主要检验样本为血、尿、脊髓液等,主要为分泌性免疫球蛋白。
抗体的“类”和“型”:抗体的类和型是不一样的,不通用,重链决定“类”,轻链决定“型”
类:决定Ig不同类的抗原性差异存在于重链的恒定区(CH)。根据CH抗原性的差异,即氨基酸组成、排列、空间构型、二硫键数目的不同,将H链分为μ、γ、α、δ、ε链五类,与L链组成完整的Ig分子。亚类主要取决于铰链区氨基酸组成和二硫键数目的差异。
型:决定Ig型的抗原性差异决定于L链的恒定区(CL)的氨基酸组成、排列和空间构型的不同,分为κ和λ两型。亚型,按λ轻链恒定区个别氨基酸的差异又可分为λ1、2、3、4四个亚型。
抗体的抗原性:同种型、同种异型和独特型
同种型:用Ig免疫异种动物将诱导产生针对该Ig的同种型抗体。同一种属内所有健康个体所共有的抗原性标志,主要集中在Ig的恒定区。间接法检测IgG使用的抗-γ链抗体和捕获法使用的抗-μ链抗体、以及各种二抗的应用,都是基于抗体的同种型抗原性。
同种异型:用Ig免疫同一种属不同个体的动物通常会诱导产生针对该Ig的抗体,即同种异型抗体。同种异型的产生是由于不同个体间Ig分子恒定区少数几个氨基酸的不同引起的,可以稳定遗传。
独特型:Ig V区上的抗原特异性,可在异种、同种异体以及自身体内诱导产生相应的抗体,称为抗独特型抗体。在Ig形成的过程中会产生各种各样的V区基因组合,在抗体亲和力成熟过程中也会使可变区发生种种变化,成为Ig独特的抗原特异性标志。
抗体的结构请看下图
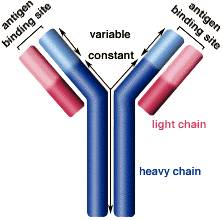
抗体的Y字形结构
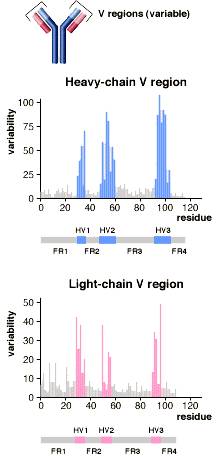
抗体可变区结构
各类免疫球蛋白的特性:
IgA:人类每天合成IgA的量大约为66mg/Kg体重,比其他所有Ig总和还多,但其半衰期只有6天,所以血液中IgA的含量少于IgG,仅占血液Ig的10%左右,但在粘膜表面和分泌液中IgA是主要的免疫球蛋白。人类IgA单体分子量为160kDa,含IgA1和IgA2两种亚类,其中IgA2还有IgA2m(1)和IgA2m(2)两个同种异型。IgA1和IgA2重链仅仅相差22个氨基酸,它们结构上的主要差别是IgA2在铰链区缺少富含O糖基化位点的13个氨基酸。
IgA可以是单体形式,也可在J链的帮助下形成二聚体、三聚体和四聚体。分泌型IgA都是多聚体。血液和粘膜组织的IgA不同。血液中的IgA以单体为主,IgA1与IgA2的比例为10:1,由在淋巴结活化并分化后进入骨髓的浆细胞合成。肠黏膜组织产生的IgA大多数为含J链的二聚体,由黏膜固有层的浆细胞合成,每天可以产生5g左右,其中IgA1与IgA2的比例为2:3.所生成的IgA大部分被pIgR运送至黏膜表面,还有一部分经门静脉入血,进入肝脏后被肝细胞表达的pIgR运至胆管,再运送至小肠。IgA可以起中和抗体的作用。
IgD:人血清IgD含量为3~40μg/mL,半衰期约2.8天。IgD是分子量为184kDa的糖蛋白,糖基占其分子量的9%,IgD只有单体形式,重链由一个可变区和三个恒定区结构域构成,由于存在一个较长的铰链区,所以易受酶降解。
IgD是B细胞发育分化成熟的标志,许多B细胞都共表达膜IgD和膜IgM,当B细胞转变为记忆细胞后,膜IgD的表达量下降直至消失。
IgE:主要由呼吸道和肠道上皮下的浆细胞分泌,在血液中IgE的含量很低,正常人血液中IgE含量约50ng/mL,血液中IgE的半衰期只有两天,过敏患者IgE的水平可以升高数倍。体内大部分IgE与组织细胞结合,细胞结合的IgE的半衰期可以延长至数个星期。
IgE的分子量为190kDa,仅有单体形式,其重链由4个恒定区。IgE的糖基化程度很高,糖基占其分子量的13%,由7个潜在的糖基化位点。在病理条件下,一些人会对抗原产生异常的高反应性,导至I型超敏反应的发生。
IgG:IgG在体内分布广泛,是血液和组织液的主要免疫球蛋白。人血液IgG1含量9mg/mL、IgG2含量3mg/mL、IgG3含量1mg/mL、IgG4含量0.5mg/mL。IgG1、2、4分子量为146kDa,半衰期约3周;IgG3分子量为165kDa,半衰期仅一周。
IgG主要在B细胞经过亲和力成熟的再次免疫应答中产生,一般来讲IgG对抗原的亲和力要比IgM高很多,是再次免疫应答的主要抗体。
小鼠IgG分为IgG1、IgG2a、IgG2b和IgG3四类。
IgM:IgM单体分子量为180kDa,含糖量为10%-12%。IgM每条重链含有4个恒定区,由于缺少铰链区,IgM分子柔韧性较差。IgM单体主要以膜蛋白的形式与IgD一起在初始B细胞上表达。
血液中IgM浓度为1.5mg/mL,大多是分子量为970kDa的五聚体,此外还有少量四聚体和六聚体。五聚体的IgM通过J链以及单体与单体之间的二硫键连接而成。
机体中的IgM主要有两个来源,一个是在B-2细胞对外来抗原的初次免疫应答中产生。虽然IgM在再次感染中也生成,但不是主要的免疫球蛋白。IgM的另一个来源是B-1细胞在病毒、细菌的诱导下经多克隆活化而产生的,它们针对的经常是病原体表面的糖成分。
机体初次免疫应答时最初产生的抗体总是IgM,此时由于B细胞的体细胞突变尚未发生,形成的IgM的亲和力较低。不过IgM是五聚体,有10个抗原结合位置(实际为五价)具有较高的亲合力。
附:机体免疫应答的三个阶段
机体对感染物的应答包括三个阶段:
固有免疫期(0-4h):当病原微生物或外来异物进入机体后,主要参与免疫系统识别和清楚病原体的是一些现存的效应分子,当感染因子不能被清除,进入固有免疫应答期。
固有免疫应答期(4-96h):固有免疫应答期是早期诱导性应答,发生在细菌入侵4-96小时,吞噬细胞识别病原体并活化,吞噬清除病原体;分泌一系列细胞因子,引起炎症反应;激活NK细胞等多种固有免疫的效应细胞,共同行使对感染引子的清除。当感染因子扔不能被清除,进入适应性免疫应答期。B1细胞可以识别并结合某些细菌表面共有的多糖抗原而被活化,在较短的时间内(48h)即可以产生低亲和力IgM为主的抗体。
适应性免疫应答期(>96h):适应性免疫应答期是晚期诱导性应答,发生在细菌入侵的96小时,未被清除的感染因子提呈给T细胞启动特异性T细胞免疫应答,B细胞直接识别抗原后诱导特异性体液免疫应答。每个B细胞开始时一般均表达IgM,在免疫应答中首先分泌IgM,但随后可表达产生IgG、IgA或IgE。影响Ig类别转换的因素主要有①抗原种类:B细胞对TI-1抗原的应答不引起抗体类型转换,一般只诱导产生IgM类抗体;对TI-2抗原的应答主要产生IgM,也可产生IgD。对TD抗原的应答则能发生类型转换。一般而言,病毒及细菌主要诱导抗体向IgG转换;寄生虫及变应原主要诱导抗体向IgE转换。②Th细胞的辅助作用,Th细胞分泌的细胞因子对Ig的类型转换也起着重要的调节作用。③与B细胞所处的解剖位置有关。




 首页
首页