上海第二军医大学长征医院 廖万清 李秀丽
现在机会性深部真菌感染的发病率急剧上升,你知道主要有哪些因素吗?
自80年代以来,深部真菌感染的发病率在逐年上升,尤其是免疫抑制等高危患者的增加,以及耐药真菌和新出现真菌的日益普遍,显著改变了深部真菌感染的流行情况,如曲霉菌及其他霉菌的感染不断增加,非白念珠菌成为念珠菌感染的主要原因。使得人们迫切需要对深部真菌感染的致病机理及对抗药物进行更进一步的研究。
深部真菌感染已经严重威胁人类健康
深部真菌感染指病原性真菌侵犯皮下组织、粘膜和/或内脏器官引起的真菌感染性疾病。近年来,由于免疫受损人群,包括恶性肿瘤、恶性血液病、艾滋病、糖尿病、自身免疫病、严重烧伤和创伤、长期吸毒以及人口老龄化等发病快速扩大,以及广谱抗生素、皮质类固醇激素和免疫抑制剂的广泛使用,导管插管和器官移植等新技术的开展,使机会性深部真菌感染的发病率急剧上升。例如有报道器官移植受者中发生深部真菌感染者:肾移植后14%,心、肺移植后35%,肝移植后42%,约 30%的急性白血病患者发生侵袭性真菌感染。在美国,近十年来住院病人的深部真菌感染率己提高了2倍;而在欧洲,在对各种死亡病人所做的尸体解剖中发现,从1978~1982年深部真菌感染率为2.2%~3.2%,1983~1987年感染率为 5.1%,而最近十年的感染高峰己达7.3%。深部真菌感染危险性也明显增高。据文献报道,近年来国内外深部真菌感染的发病率明显增多,其病死率可高达29%~60%。在有严重基础疾病的患者预后严重,病死率高,例如念珠病病死率可达40%,侵袭性曲霉病可高达50%~100% 。深部真菌病的主要病原菌包括念珠菌、隐球菌、曲霉菌、孢子丝菌、着色霉菌、毛霉、马内菲青霉、组织胞浆菌、副球孢子菌和皮炎芽生菌等,其中念珠菌病、隐球菌病和侵袭性曲霉病最常见。美国一份报道显示,与80年代相比,念珠菌病的年发病率由26%增至72.8%,隐球菌病由4.0%增至6.55%, 曲霉病由8.4%增至12.42 % 。因此,常见深部病原真菌的致病机理的研究十分活跃,现以白念珠菌、烟曲霉、新生隐球菌为例,就这方面内容加以总结。
常见深部病原真菌的研究
(一)念珠菌以白念珠菌为主要致病菌,其致病主要与粘附能力、受体 、形态转换、胞外酶等有关。⒈粘附能力与受体 白念珠菌致病的首要条件是其粘附性。其粘附物质有细胞壁上的甘露聚糖(Mn)或甘露聚糖-蛋白质复合物(M-P)、几丁质、受体和粘附素。白念珠菌Mn或MP为细胞壁主要成分,它可影响和干扰与炎症密切相关的细胞因子。白念珠菌粘附组织细胞主要通过Mn或MP与宿主细胞表面受体相互作用实现,粘附能力强的白念珠菌有较高的疏水性,较亲水性白念珠菌粘附的细胞范围广泛。研究者认为,菌体表面的疏水分子即为白念珠菌孢子的壁蛋白和菌丝壁 MP。
几丁质与白念珠菌的毒力关系密切。几丁质合成酶缺陷突变株,感染小鼠后死亡率较无缺陷株的死亡率低。用几丁质合成酶抑制剂多氧霉素D作用白念珠菌后,此菌的粘附能力明显降低。
人们发现致病性白念珠菌细胞壁上有C3b和iC3b受体,非致病性白念珠菌则无此受体。这两种受体与人类吞噬细胞膜上的相应受体有类似作用,它们可与人的免疫细胞竟争补体,从而抑制补体的调理作用,有利于白念珠菌逃避吞噬作用 。白念珠菌中存在着一个粘附素家族,有凝集素样粘附素1(ALAI),凝集素样序列1(ALSI),菌丝孔蛋白1(HWPI)及整联蛋白巫(INFI)等编码基因编码的糖蛋白。研究发现ALAI介导的粘附步骤,首先是白念珠菌迅速附着于细胞外基质蛋白上,然后细胞与细胞相互作用为主的一个相对缓慢的过程[11]。HWPI的粘附方式与ALAI不同, HWPI能与颊上皮细胞的未确定的蛋白质交联。这两种方式可组合成一个二阶段模式的粘第一阶段由传统的粘附素将芽生孢子固定在受损的上皮细胞、内皮细胞或血纤蛋白袖形血栓上,使芽管形成;第二阶段,当宿主的防御能力降低时,真菌生长旺盛,一旦局部微环境中含有了宿主TGase,即由非传统的粘附素不可逆地附着于宿主蛋白质上[12]。⒉白念珠菌的致病力与形态转换有关。白念珠菌能以三种形态生长,即菌体形态、假菌丝形态及菌丝形态。假菌丝状态可产生芽管特异性抗原,此抗原能明显增强其粘附性,能阻止中性粒细胞的吞噬杀伤作用。白念珠菌被吞噬后,在吞噬细胞内仍能生成芽管并长出菌丝,穿透宿主细胞膜。实验表明,以菌丝形态生长的菌株具有更强的致病性。MAPK(mitogen activated proemkinase,促有丝分裂原活化的蛋白激酶)信号转导途径是白念珠菌形态转换过程的途径之一[13]。cAMP途径也影响白念珠菌的形态转换。此外,尚有其他途径,如tupl基因、CRKI基因、PHRI基因,与白念珠菌的形态转换有关,对白念珠菌的形态变化调控机理的研究正在不断深入。⒊胞外酶 白念珠菌能产生水解酶、脂酶及蛋白酶等多种酶类,其中以分泌型酸性蛋白酶(SAP,又称胞外天冬氨酸酶)的研究最多。分泌SAP的毒性作用已得到充分肯定,它参与白念珠菌破坏组织。已发现4种分泌型酸性蛋白酶同功酶,分别是不同位点的基因编码产物。SAP是大多数临床白念珠菌分离物的SAP酶谱中的主要同功酶。SAP2P有分解粘蛋白的特性。目前发现SAP至少由一个多基因家族的9个基因成员编码[14]。SAP4一SAP6可在酵母-菌丝转化中表达。白念珠菌产生的磷脂酶亦与该菌侵袭力有关。
(二)烟曲霉
近年来,曲霉病的发病率明显增多,烟曲霉己成为临床上仅次于白念珠菌的一种重要的条件致病真菌,其某些成分或毒素可直接造成机体损害,或降低机体防御能力而有利于曲毒感染形成或扩散,致侵袭性曲霉病,严重可危及生命,井非所有曲霉均致病,仅少数菌株感染人体,这与少数菌株能分泌某些致病因子有关,为此,有关烟曲霉相关的致病因子及作用机制研究十分活跃,现就这方面内容加以总结。
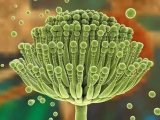
烟曲霉菌分生孢子头1000倍
目前已研究发现,烟曲霉可产生一些真菌毒素如烟曲霉素、粘帚霉毒素和烟曲霉酸以及内毒素、C物质等[15],上述这些毒性物质,可通过抑制免疫反应,或破坏局部组织,而促进真菌生长繁殖。
可产生多种细胞外酶,包括核酶、磷酸酶、肽酶和蛋白酶,这些酶可以降解大分子物质,为真菌生长提供营养,其中蛋白酶研究较为深入。临床病变部位分离的烟曲霉所产生的蛋白酶活性,比非临床分离菌株要强,说明不同菌株侵袭力差别与烟曲霉蛋白溶解作用强弱有关[15]。烟曲霉所产生蛋白酶可以促进真菌与宿主组织粘附和穿透。
你知道吗?烟曲霉是通过哪些方式侵犯肌体的?
⒈扰乱粘膜防御功能烟曲霉感染常有基础疾病,基础疾病破坏了正常呼吸道防御体系,利于烟曲霉感染形成。烟曲霉培养上清液可降低纤毛运动和损害呼吸道上皮细胞,其中以粘帚霉毒素最为重要,烟曲霉素、烟曲霉酸也参与这一损害过程。损害纤毛运动有利于烟曲霉定植,而粘膜上皮细胞损害不仅有利于烟曲霉侵入,也可以通过损害的上皮细胞释放营养物质,促进真菌繁殖[16]。⒉抑制吞噬细胞功能烟曲霉多种代谢物可干扰吞噬细胞功能,较为重要的有烟曲霉素,AFALP和局限曲霉素。⒊降低调理作用通过某些调理素增强真菌与微生物间粘附,也是机体抗真菌感染重要机制之一。真菌也可通过抑制补体激活或粘附能力,来降低吞噬细胞调理作用。⒋促进曲霉与组织粘附曲霉与组织粘附是发生曲霉病的重要步骤。烟曲霉与细胞外基质和血清蛋白粘附的成分有层连蛋白、纤连蛋白、胶原、原胶原和补体C3 等,其中尤以层连蛋白研究较,可促进曲霉与组织粘附。 (三)隐球菌隐球菌有17属和9个变种,而新生隐球菌是其主要致病菌。
⒈新生隐球菌首先进入肺泡间质
为什么说担孢子是隐球菌病的感染繁殖体,你能想到哪些支持点?
新生隐球菌感染可能初发于机体吸入担孢子。理论上,能穿过肺组织的最大颗粒直径为2m,担孢子的直径为1.8~2m,并且对干燥有抵抗力。Wickes等[17]发现新生隐球菌在异宗配合的条件下可以产生菌丝和担孢子,这个过程与交配型有关。新生隐球菌单倍体的交配型细胞在合适条件下随着孢子和有生育能力的担孢子的出现可形成单核菌丝。来自环境和临床的分离标本,这两种交配型的比例分别是40:1和30:1。交配类型的优势性,担孢子抵抗干燥的能力和合适的大小等特点支持担孢子是隐球菌病的感染繁殖体。⒉新生隐球菌在宿主体内生存的机制新生隐球菌进入宿主体内,必须适应36~37℃的生长条件,这种转变需要特异的生长因子。Odom等[18]证实钙神经素是新生隐球菌在37℃生长所需要的生长因子。 感染初期,大多数新生隐球菌对非活化的吞噬细胞有一定抗吞噬性,荚膜合成是一种主要的抵抗吞噬的保护机制[19]。此外细胞介导的免疫力直接参与宿主的防御反应。用新生隐球菌培养滤过抗原免疫小鼠,可以诱发两种淋巴细胞群。其中Tamp淋巴细胞能产生大量Th1相关淋巴因子,通过激活巨噬细胞和可能的其他抗隐球菌的效应细胞提供免疫保护[20]。白细胞介素12(IL-12)、-干扰素亦可抑制新生隐球菌的活力[21]。抗体的保护作用集中于对抗新生隐球菌葡萄糖醛酸甘露聚糖系列单克隆抗体的研究。此单抗存在时,新生隐球菌与人单核细胞和T细胞共同孵育可以提高-干扰素、IL-1和IL-12的产生, 而对IL-10的产生是下调作用[22]。这促使特异单克隆抗体调节细胞因子的表达和T细胞增殖。据此人们推测特异抗体通过促进Th1相关细胞因子的反应而具保护作用。
⒊新生隐球菌逃逸宿主防御反应的机制当感染免疫正常的宿主时,这些机制是无效的;但对免疫缺陷的宿主,隐球菌可以有效地逃逸防御反应,引发播散感染。Varki等[23]推测在感染初期荚膜多糖完全表达之前,硅酸可能是新生隐球菌抗吞噬的重要成分。有通过体外实验表明,无荚膜酵母细胞比产荚膜的菌株或加了多聚糖的无荚膜株更易被吞噬,遗传学研究也支持荚膜的产生与其毒性相关[24]。Chaturvedi等[25]证实在培养液以及感染的兔模型中,新生隐球菌产生大量的甘露醇。人们推测甘露醇通过取游离的氢氧分子,保护新生隐球菌免受吞噬细胞的氧化性杀伤。新生隐球菌产生依赖铜离子的漆酶,该酶可以催化黑素样色素的形成。人们已经证实了编码漆酶结构基因的CNLAC1基因在新生隐球菌的毒性中的作用[26]。⒋ 最近,新生隐球菌的信号传导研究取得了明显的进展。温度、荚膜及黑素三种毒性表型的信号传导路径已经确定。许多真菌与新生隐球菌所采用这种路径的差异性已经被阐明。深部真菌感染研究展望以上以最常见的几种深部致病真菌为例,对深部真菌的致病机制进行了初步的阐述。随着免疫受损人群发病快速扩大,以及广谱抗生素、皮质类固醇激素、免疫抑制剂的广泛使用,器官移植等新技术的开展,使机会性深部真菌感染的发病率急剧上升。许多条件致病菌致病报道明显增多,有孢子丝菌病、着色芽生菌病、毛霉病、蛙粪霉病、马内菲青霉病、地霉病、组织脑浆菌病、球袍子菌病、副球袍子菌病、皮炎芽生菌病等,如赵俊英等报道了天疱疮合并肺部奔马赭霉暗色丝孢霉病一例。高爱平等报道了颈囊虫病合并囊内申克氏真菌感染一例,廖小梅等报道了艾滋病合并播散性马尔尼菲青霉菌感染一例,我科报道了萨瓦曲霉引起肺感染一例等等。随着现代医学的迅速发展,先进诊疗技术的引进,临床深部真菌感染日益增多且趋向复杂化,常导致致死性终末感染,对深部真菌致病机制研究的不断深入,可为开发深部真菌病的早期诊断奠定基础,也为寻找防治深部真菌感染的提供途径和措施,为研制疫苗,开发新的药物提供了理论依据。致病基因的克隆,在深部真菌的基因诊断和分类鉴定等方面也将得到广泛应用。对深部真菌致病机制的研究,任重道远,路漫漫其修远兮,需吾等上下求索。
小结
机会性深部真菌感染的发病率急剧上升;
介绍了白念珠菌、烟曲霉、新生隐球菌深部病原真菌的致病机理。
 首页
首页