电压门控钠(Nav)通道通过启动和传播动作电位来控制膜的兴奋性。与它们的生理意义一致,这些通道的功能障碍或突变与各种通道病有关。因此,Nav通道是各种临床和研究药物的主要靶点。此外,大量的天然毒素,包括小分子和多肽,都可以与Nav通道结合并调节其功能。冷冻电子显微镜(cryo-EM)的技术突破已经能够确定真核生物(最终是人类)Nav通道的高分辨率结构,无论是单独的还是与辅助亚基、毒素和药物的复合。这些研究不仅提高了我们对通道结构和工作机制的理解,而且为原型药物和毒素的结合和作用机制(MOA)的分子基础提供了前所未有的清晰度。
2023年11月30日,清华大学/深圳医学科学院颜宁团队在Channels 在线发表题为“A structural atlas of druggable sites on Nav channels”的综述文章,该综述将概述Nav通道结构药理学的最新进展,包括配体结合Nav通道的结构图谱。这些发现为未来的药物开发奠定了重要的基础。

电压门控钠(Nav)通道是一种跨膜蛋白,在膜去极化时选择性地输入钠离子。Nav通道对于可兴奋细胞(如神经元和肌肉细胞)的电信号传递至关重要,并负责动作电位的产生和传播。钠电流的电压依赖性激活最早由Hodgkin和Huxley在近70年前记录,标志着动作电位的发现。Hille研究了Nav通道的离子选择性,并提出了“四屏障,三位点模型”来描述钠离子的选择性。他还分析了局麻药与相关Nav靶向药物的结合模式。Armstrong和Bezanilla使用高分辨率电生理记录方法测量门控电流,并探索了Nav通道快速失活的过程和决定因素。在20世纪80年代初,Barchi和Catterall小组研究了Nav通道的生化特性;Numa等报道了Nav通道成孔亚基(α亚基)的编码序列。总之,这些里程碑建立了Nav通道的功能、工作机制和结构特性的早期概念。
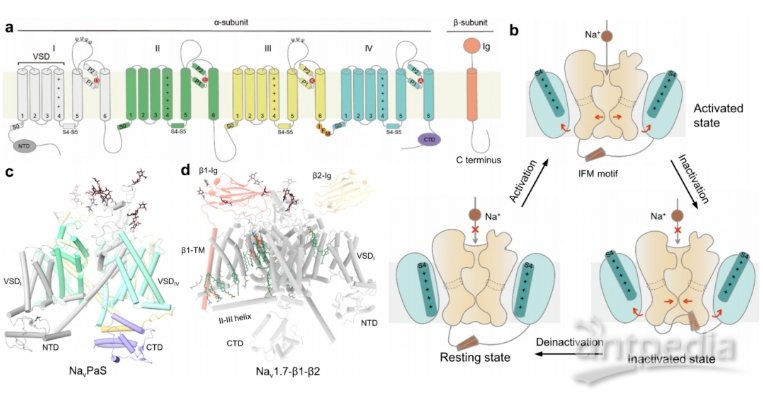
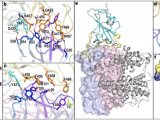
Nav通道属于电压门控离子通道(VGIC)超家族。它们通常包括一个核心α亚基,其本身足以进行通道活动,以及辅助β亚基(β1-β4),有助于膜定位和通道调节。α亚基是一个长度约为2000个残基的单一多肽,折叠成4个同源重复序列(重复I-IV)。每个重复序列由6个跨膜螺旋组成(S1-S6),其中四个重复序列中的S5和S6构成离子渗透孔域(PD),每个重复序列中的S1-S4构成侧翼电压感应域(VSD)。S5和S6之间的序列组装成细胞外环(ecl)和离子选择性过滤器(SF)。四个不同的残基,Asp/Glu/Lys/Ala (DEKA) ,位于每个重复的相应SF位点,负责钠离子选择性。III-IV连接体包含Ile/Phe/Met (IFM)基序,这对于Nav通道的快速失活至关重要。这种疏水簇的突变可以完全消除这种快速失活。
Nav通道的结构和一般工作机制概述(图源自Channels )
人类Nav α亚基有9个亚型,分别为Nav1.1-Nav1.9。Nav1.1-Nav1.3和Nav1.6主要在中枢神经系统表达。Nav1.4和Nav1.5分别存在于骨骼肌和心肌中。Nav1.7-Nav1.9主要作用于周围神经系统。
Nav通道是产生膜兴奋性的关键。即使是电压依赖性钠电流的微小变化也会产生有害影响。Nav通道的功能障碍或异常调节与多种疾病有关,主要与通道的组织特异性有关。例如,在Nav1.1的编码基因SCN1A中,除了数百个非义突变外,还发现了许多错义突变,使其成为癫痫发作的最易感危险因素。Nav1.5是致病突变的另一个热点。在心脏疾病患者中发现了数百种突变,如3型长QT综合征、Brugada综合征和纤颤。包括Nav1.3和Nav1.6-Nav1.9在内的几种Nav通道亚型与痛觉有关。Nav通道一直是治疗癫痫、心律失常、精神障碍和疼痛障碍的主要药物靶点。
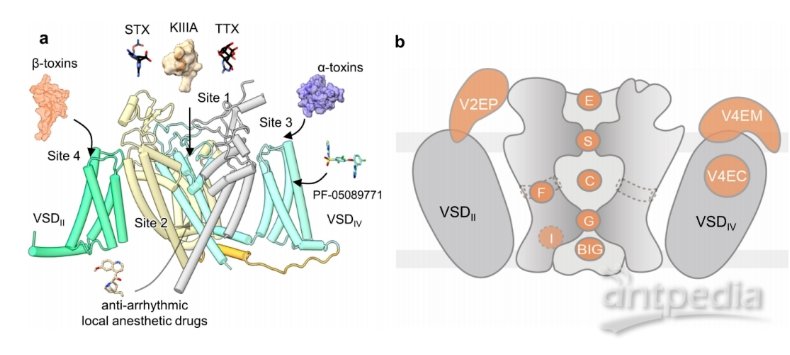
Nav通道上配体结合位点的结构作图(图源自Channels )
剖析Nav通道的工作和发病机制以及Nav靶向药物的作用模式(MOA)需要不同功能状态下的高分辨率结构。Nav通道的简化工作图包括三种主要状态:休眠、激活和未激活。在静息膜电位下,Nav通道不导电,PD闭合,VSD呈“向下”构象。作为对膜去极化的响应,S4螺旋被驱动到细胞外侧以实现“上”构象。VSD的运动导致PD的打开,这一过程被称为机电耦合。激活后,通道在1 ~ 2 ms内迅速失活。在膜超极化时,Nav通道从失活状态释放到静息状态,为下一个工作周期做好准备。
冷冻电子显微镜(cryo-EM)技术和算法的突破为近原子分辨率的Nav通道可视化提供了前所未有的机会。2017年发表了真核生物Nav通道的第一个结构,即以美洲大蠊(Periplaneta americana)的短亚型命名的NavPaS。几个月后,研究人员报道了从电鳗 (EeNav1.4)中分离出的β1结合的Nav通道的冷冻电镜结构。这些研究揭示了不同于同四聚体VGIC的单链Nav通道的组装原理和结构细节。出乎意料的是,这两个结构在完全不同的状态下被捕获。NavPa具有收缩的孔域结构,4个VSD呈明显的向上构象,而EeNav1.4的孔域结构在胞内门处松弛,4个VSD均呈类似的“向上”状态。最显著的结构差异发生在短的III-IV连接体上。基于结构差异,研究人员提出了IFM基序快速失活Nav通道的“门楔”变构阻断机制。
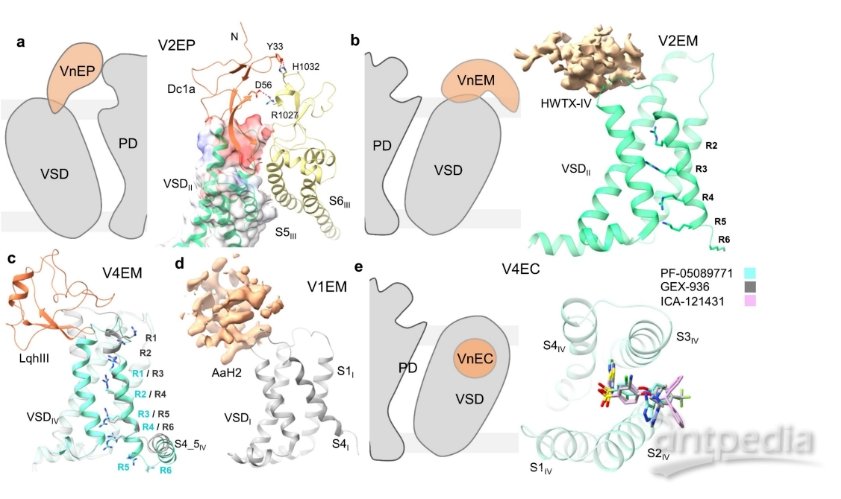
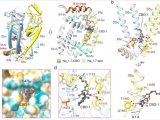
VSD上的结合位点(图源自Channels )
在随后的几年里,研究人员最终捕获了人类Nav1.1、Nav1.2和Nav1.4-Nav1.8的结构,其中大部分与辅助亚基和毒素/药物复合物。Catterall和Jiang组分别报道了大鼠Nav1.5 (rNav1.5)和人Nav1.3的结构。这些结构共同为理解过去半个世纪积累的大量实验和临床数据提供了分子基础。在这篇综述中,将重点介绍Nav通道的结构药理学的最新进展,这对未来合理的药物开发提供了重要的见解。












































































































































































































































































































 首页
首页

