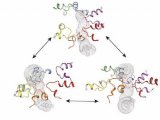
mTORC1可调节细胞生长和代谢以响应多种营养素,包括必需氨基酸亮氨酸。最近在培养的哺乳动物细胞中的工作将 Sestrins 确定为亮氨酸结合蛋白,在亮氨酸剥夺期间抑制 mTORC1 信号传导,但它们在机体对膳食亮氨酸的反应中的作用仍然难以捉摸。
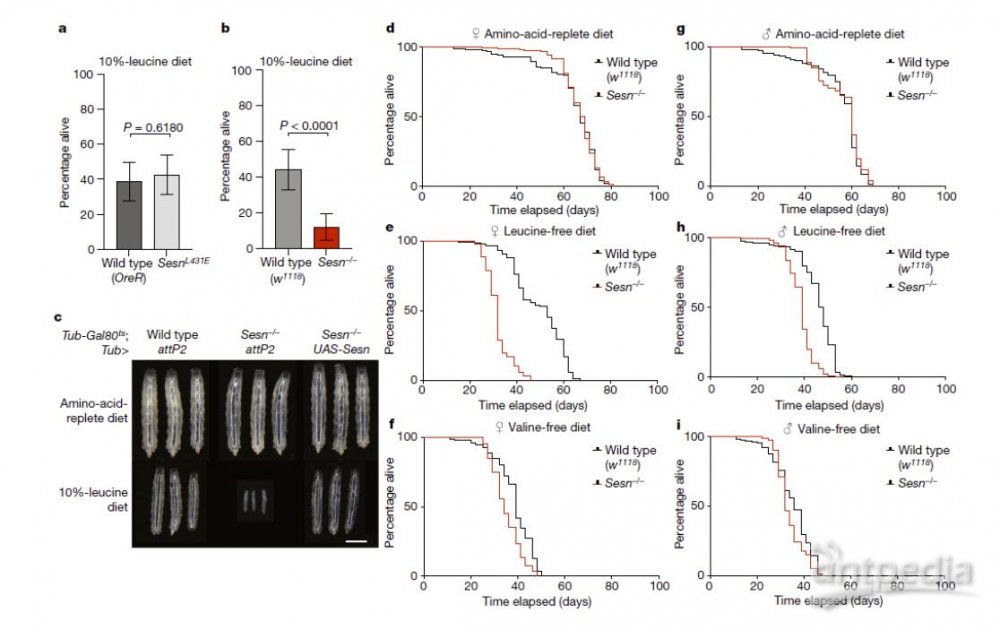
2022年7月20日,David M. Sabatini团队(Sabatini没有任何附属单位)(麻省理工学院Xin Gu,哈佛医学院Patrick Jouandin及Norbert Perrimon为共同通讯作者)在Nature 在线发表题为“Sestrin mediates detection of and adaptation to low-leucine diets in Drosophila”的研究论文,该研究发现 Sestrin-null 果蝇 (Sesn−/−) 在急性亮氨酸饥饿后无法抑制 mTORC1 或激活自噬,并且在低亮氨酸饮食中发育受损和寿命缩短。表达亮氨酸结合缺陷型 Sestrin 突变体 (SesnL431E) 的敲入果蝇降低了对亮氨酸不敏感的 mTORC1 活性。
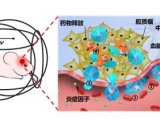
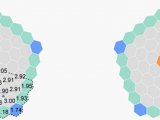
值得注意的是,该研究发现果蝇可以区分含或不含亮氨酸的食物,并优先以含亮氨酸的食物为食并产下后代。这种偏好取决于 Sestrin 及其结合亮氨酸的能力。亮氨酸调节神经胶质细胞中的 mTORC1 活性,而这些细胞中 Sesn 的敲低会降低果蝇检测无亮氨酸食物的能力。因此,mTORC1 的营养感应对于果蝇来说不仅是为了适应,而且是为了检测缺乏必需营养素的饮食。
另外,2021年12月2日,麻省理工学院David Sabatini等团队(注:由于David M. Sabatini 不再隶属于怀特黑德研究所或霍华德休斯医学研究所, 为确保执行通讯作者的职责,Jessica B. Spinelli 作为通讯作者)在Science 在线发表题为“Fumarate is a terminal electron acceptor in the mammalian electron transport chain”的研究论文,该研究发现当氧还原受阻时,复合物 I 和二氢乳清酸脱氢酶 (DHODH) 仍然可以将电子沉积到电子传递链 (ETC)中。缺乏氧还原的细胞积累泛醇,反向驱动琥珀酸脱氢酶 (SDH) 复合物,使电子沉积到延胡索酸上。在抑制氧还原后,延胡索酸还原维持 DHODH 和复合物 I 的活性。 小鼠组织显示出使用延胡索酸盐作为终端电子受体 (TEA)的不同能力,其中大部分在缺氧条件下会逆转 SDH 复合物。因此,该研究描绘了哺乳动物 ETC 中的电子流回路,该回路在氧限制下维持线粒体功能。
另外,近期David Sabatini深陷性骚扰风波,MIT怀特黑德生物医学研究所发布正式声明,宣布David Sabatini不再与怀特黑德研究所及霍华德休斯医学院存在任何关联,并在麻省理工学院启动自己的调查期间休了行政假。

蛋白激酶 mTORC1 响应多种信号调节生长和新陈代谢,包括生长因子和氨基酸等营养物质。氨基酸通过促进 mTORC1 易位到溶酶体表面来激活 mTORC1,这是其必需的激活剂 Rheb 所在的位置。异二聚体 Rag GTPases 受多种多组分蛋白质复合物(包括 GATOR1 和 GATOR2)控制,调节 mTORC1 的溶酶体定位。
GATOR1 是 RagA 和 RagB 的 GTPase 激活蛋白,是氨基酸剥夺以抑制 mTORC1 信号传导所必需的。相比之下,GATOR2 是氨基酸激活 mTORC1 所必需的,并直接与迄今为止发现的几种氨基酸感应器相互作用,这表明尽管其生化功能仍未知,但它仍可作为营养感应中心。
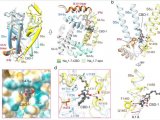
在蛋白质氨基酸中,亮氨酸是最成熟的 mTORC1 激活剂。在培养的哺乳动物细胞中的研究表明,亮氨酸通过调节 GATOR2 与 Sestrin 蛋白家族的相互作用来控制 mTORC1,这些蛋白是 mTORC1 信号传导的阻遏物。人 Sestrin1 和 Sestrin2 结合亮氨酸的亲和力与激活 mTORC1 所需的亮氨酸浓度一致,并且是剥夺亮氨酸以抑制 mTORC1 信号传导所必需的。
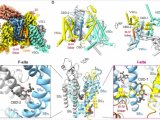
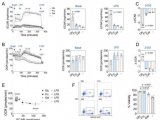
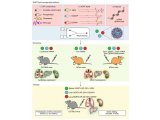
果蝇需要 Sestrin 来适应低亮氨酸饮食(图源自Nature )
此外,不结合亮氨酸的 Sestrin2 突变体在亮氨酸存在下无法与 GATOR2 分离,并且在表达该突变体的细胞中,即使在亮氨酸充足的条件下培养细胞时,mTORC1 活性仍然很低。尽管有证据表明 Sestrin 是培养的哺乳动物细胞中 mTORC1 通路的亮氨酸感应器,但 Sestrin 介导的亮氨酸感应在完整生物体生理学中的作用在很大程度上仍未得到探索。
尽管亮氨酸感应的大部分工作是在哺乳动物系统中进行的,但 Sestrin 和核心营养感应机制,包括 Rag GTPase、GATOR1 和 GATOR2,在大多数无脊椎动物中都是保守的,包括黑腹果蝇。与哺乳动物不同,果蝇仅表达一个 Sestrin (Sesn) 基因,极大地促进了 mTORC1 对亮氨酸传感的体内研究。
该研究发现 Sestrin-null 果蝇 (Sesn−/− ) 在急性亮氨酸饥饿后无法抑制 mTORC1 或激活自噬,并且在低亮氨酸饮食中发育受损和寿命缩短。表达亮氨酸结合缺陷型 Sestrin突变体 (SesnL431E) 的敲入果蝇降低了对亮氨酸不敏感的 mTORC1 活性。
值得注意的是,该研究发现果蝇可以区分含或不含亮氨酸的食物,并优先以含亮氨酸的食物为食并产下后代。这种偏好取决于 Sestrin 及其结合亮氨酸的能力。亮氨酸调节神经胶质细胞中的 mTORC1 活性,而这些细胞中 Sesn 的敲低会降低果蝇检测无亮氨酸食物的能力。因此,mTORC1 的营养感应对于果蝇来说不仅是为了适应,而且是为了检测缺乏必需营养素的饮食。







































































































































































































































































 首页
首页


